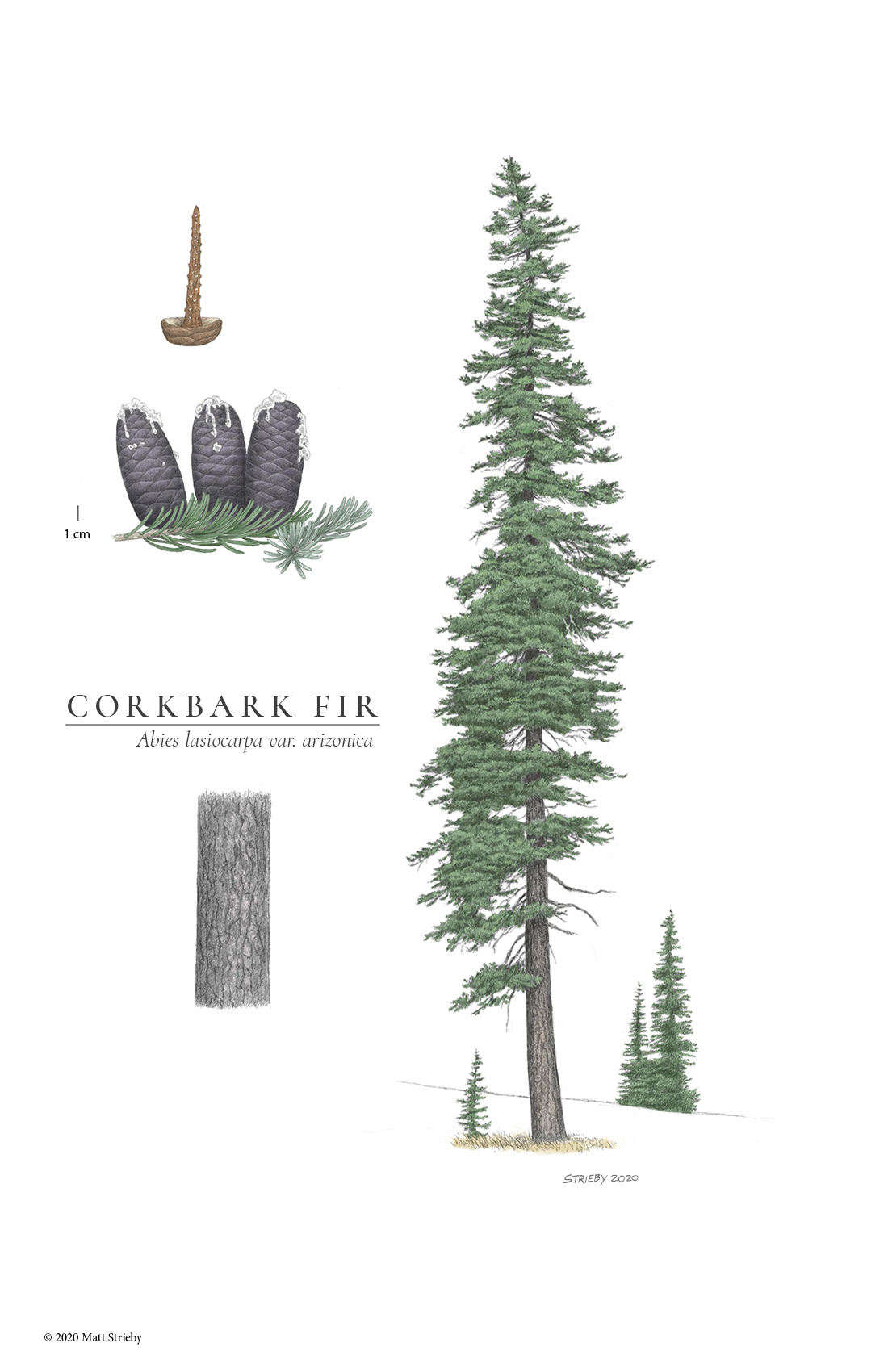
Drawing of var. arizonica by Matt Strieby [2020].

Mature tree about 50 cm dbh and 25 m tall, Entiat River valley, Washington [C.J. Earle, 2004.07.02].

Mature seed cones, Rocky Mountain National Park, Colorado [C.J. Earle, 2006.07.04].

Fully mature seed cones, some of which are starting to disintegrate, on a tree near Hyas Lake, North Cascades, Washington [C.J. Earle, 2003.08.23].

Scales and seeds from disintegrating cones floating on a pond near Mount Adams, Washington [C.J. Earle, 2009.09.26].

Ripe pollen cones on a tree at Manastash Ridge, Washington [C.J. Earle, 2015.05.23].

Foliage, Mount Rainier, Washington. Current growth is much more glaucous than older foliage [C.J. Earle, 2002.09.07].

Typical shade foliage, including new growth [C.J. Earle, 2016.06.18].

Twig with bark peeled back to show the red periderm that identifies var. lasiocarpa. Width of image 1 cm. Collected at Sunrise, Mount Rainier [K. Hagmann, 2008.08].

Bark of a corkbark fir (var. arizonica) growing on Mount Lemmon, Santa Catalina Mountains, Arizona [Jeff Bisbee].

Bark on a roadside tree of var. bifolia 50 cm diam. [C.J. Earle, 2006.07.04].

Seedling at least 2 years old (no cotyledons), Yellowstone National Park [C.J. Earle, 2019.08.22].

Dwarf mistletoe covered in ripe seeds, on var. bifolia [C.J. Earle, 2007.07.22].

Tree clump, eastern Olympic Mountains, Washington. Multiple stems layered from a parent plant that either no longer exists or is of small stature. Areas barren of foliage reflect past frost kill of portions exposed above the winter snowpack [C.J. Earle, 2011.09.04].

This large tree at Galena Pass, Idaho evidently lost its top many years ago and has since regrown at least seven codominant leaders [C.J. Earle, 2012.09.29].

Tree knocked down, in which at least 18 branches have independently assumed apical dominance even though all share the same base and root system [C.J. Earle, 2018.10.13].

Conservation Status

Abies lasiocarpa
(Hooker) Nuttall 1849
Common names
Subalpine fir, alpine fir, balsam fir, white fir, mountain balsam fir, white balsam, western balsam fir, Rocky Mountain fir; pino real blanco de las sierras [Spanish] (Burns and Honkala 1990), sapin concolore (Hunt 1993). The type variety is here called northern subalpine fir. Var. bifolia is sometimes called the Rocky Mountain subalpine fir, and var. arizonica is commonly called corkbark fir or alamo de la sierra (Burns and Honkala 1990).
There are a few stodgy ecologists out there who maintain that "alpine" means above the treeline. They have a point, but I won't complain if somebody calls it "alpine fir." Besides, "subalpine" fir has enormous elevational range, occurring from sea level (in Alaska) to 3,700 meters (in Arizona), and can be found in many lowland or montane forests. The epithet "lasiocarpa" means "woolly seed" so we could call it woollyseed fir. It's closely related to Abies balsamea, so "western balsam fir" might be the best name, but it's almost never used.
Taxonomic notes
There are three distinct types of Abies lasiocarpa, which are here described as varieties:
- A. lasiocarpa var. lasiocarpa, the northern subalpine fir, described from Mount Hood, Oregon.
- A. lasiocarpa var. arizonica (Merriam) Lemmon, the corkbark fir, native to the southern Rocky Mountains.
- A. lasiocarpa var. bifolia Eckenwalder 2009, the western subalpine fir, native to much of the interior of western North America.
Although it is so named in the Flora of North America, most conifer biologists have chosen not to recognize var. bifolia as a separate species, because only very minor characters distinguish it from typical A. lasiocarpa. The variety is said to be distinct from var. lasiocarpa in chemical tests on wood, lack of crystals in the ray parenchyma, lack of lasiocarpenonol, and distinct terpene patterns. Var. bifolia also tends to have slightly shorter and fewer prominently notched leaves than var. lasiocarpa. The two are separated by the color of their periderm and by the shape of their basal bud scales (Hunt 1993). My experience, however, is that these characters are continuously varying and they can be found throughout the range in which I have encountered the species, which extends from southern British Columbia into Oregon and east to Montana, Wyoming, and Colorado. Adams et al. (2011) examined essential oils and DNA sequences in populations presumably representing the three varieties (Olympia Mountains of Washington, eastern Rocky Mountain Front in Montana, Wasatch Mountains in Utah, San Francisco Peaks in Arizona) and found only very small DNA differences but consistent essential oil differences, consistent with the notion that varietal distinctions are a function of adaptation to different climates. I doubt that even an expert can consistently distinguish vars. lasiocarpa and bifolia based solely on morphological characters. Distinguishing between var. bifolia and var. arizonica is also ambiguous, and is primarily based on the thickened, 'corky' bark of var. arizonica that may also be dark grey and deeply furrowed.
In north central Alberta, var. bifolia introgresses with A. balsamea (Hunt and von Rudloff 1974, Moss 1953, both cited by Hunt 1993). At the southern end of its range, var. lasiocarpa possibly hybridizes with A. procera, but the taxa are normally separated by habitat and elevation. Var. lasiocarpa shares with A. procera a red periderm, crystals in the ray parenchyma, and reflexed tips of the bracts, features not shared with var. bifolia (Hunt 1993). A. lasiocarpa (var. lasiocarpa and var. bifolia) and A. amabilis, although sympatric over a wide area, are separated by many morphologic features, and no hybrids have been found (Hunt 1993).
Unique populations of var. lasiocarpa from coastal Alaska are found at lower elevations (0-900 m) and appear to be isolated with no reported introgression between them and the coastal mountain populations (Heusser 1954, Harris 1965). The population on the Prince of Wales Island has distinct terpene patterns and warrants further study to clarify its separation from neighboring populations (Hunt 1993).
Synonymy:
For var. lasiocarpa, Pinus lasiocarpa Hooker 1838.
For var. bifolia, Abies bifolia A. Murray, Abies subalpina Engelmann, and A. lasiocarpa subsp. bifolia (A. Murray) Silba 2008.
Description
Trees to 50 m tall and 200 cm DBH, often much smaller due to growth in harsh environments; all varieties occur as krummholz at alpine elevations. Crown usually tall-conical, becoming somewhat flattened and irregular in old trees. Primary branches stiff, whorled at right angles to axis of a terete, monopodial trunk. Bark gray, thin, smooth, with resin blisters in young trees, becoming furrowed and scaly with age. Twigs stout, stiff, sunlit branchlets with uniform branching pattern, green-gray to light brown, sparse brown pubescence. Buds hidden by leaves or exposed, tan to dark brown, nearly globose, small, resinous, apex rounded; basal scales triangular to spathulate, glabrous or with a few trichomes at base, not resinous, margins entire dentate, apex acute to rounded. Leaves 11-31 × 1.25-2 mm, spiraled and turned upward, flexible; cross section flat, grooved on the upper surface; lower surface with 3-5 lines of stomata on each side of midrib; upper surface light green to blue-green, usually glaucous, with 3-6 rows of stomata at midleaf that are continuous to leaf base and more numerous toward leaf apex; resin canals large, ± medial; leaf apex notched to rounded. Cotyledons 3-6. Pollen cones at pollination ± purple to purple-green. Seed cones dark purple-blue to gray-purple with a rounded apex, sessile, 5-12 × 2-4 cm. Cone scales ca. 1.5 × 2.5 cm, densely pubescent; bracts included (specimens with exserted, reflexed bracts are insect infested). Seeds 5-7 × 2-3 mm, brown with a light brown wing about 1.5 times as long as the nut. 2n=24 (Hunt 1993 and pers. obs.). See García Esteban et al. (2004) for a detailed characterization of the wood anatomy.
Hunt (1993) asserts that the following characters serve to discriminate the varieties:
Var. lasiocarpa: Pulling off a few needles will show a red periderm on the fresh leaf scars (see photo at right). On foliar buds, the basal scales are equilaterally triangular with crenate or dentate margins. Leaves 18-31 mm × 1.5-2 mm; odor sharp (ß-phellandrene); lower surface with 4-5 stomatal rows on each side of midrib; upper surface blue-green, very glaucous, with 4-6 stomatal rows at midleaf.
Var. bifolia: Fresh leaf scars reveal a yellow or tan periderm. On foliar buds, the basal bud scales are long, narrow-triangular to spathulate with crenate to entire margins. Leaves 11-25 × 1.25-1.5 mm; odor similar to camphor; lower surface with 3-5 lines of stomata on each side of midrib; upper surface light green to blue-green, usually glaucous, with 3-6 rows of stomata at midleaf.
Var. arizonica: Has thickened, 'corky' bark that is white to grey and deeply furrowed, and has very glaucous foliage. In other respects it is like var. bifolia.
In general, I have found that the tall, narrow, spire-like crown tends to identify this species even at a great distance. Closer up, the smallish grey cones (usually 7-10 cm) and the sun foliage with stomata on the upper surface are usually sufficient to distinguish it from any similar species.
Distribution and Ecology
Canada: Yukon, Northwest Territories, British Columbia, Alberta; United States: Washington, Oregon, California, Idaho, Montana, Wyoming, Colorado, New Mexico, Arizona, Utah and Nevada. See also Thompson et al. (1999). Hardy to Zone 5 (cold hardiness limit between -28.8°C and -23.3°C) (Bannister and Neuner 2001).
Distribution data from USGS (1999).
Var. lasiocarpa occurs in Canada: S Yukon and the Coast Range of British Columbia; United States: The coast ranges of SE Alaska, Washington, Oregon, and California (Hunt 1993). My own observations at numerous sites in the Olympic and Cascade Mountains have indicated that most of these trees key out as var. bifolia. Typically found at 1100-2300 m elevation in coastal subalpine conifer forests (Peattie 1950), var. lasiocarpa grows to the alpine treeline in most of its range (Little 1980). Common associates include Abies amabilis, Pinus albicaulis and Tsuga mertensiana.
Var. bifolia occurs in Canada: S Yukon, Northwest Territories, Alberta, British Columbia; United States: Washington, Oregon, Idaho, Montana, Wyoming, Colorado, New Mexico, Arizona, Utah and Nevada at 600-3700 m in subalpine conifer forests (Peattie 1950, Hunt 1993). Var. bifolia is found growing to the alpine treeline in most of its range. In most of the Rocky Mountains, it forms a major forest type with Picea engelmannii (Little 1980).
Var. arizonica occurs in United States: Arizona, Colorado and New Mexico, typically at elevations of 2400-3400 m, where it commonly grows with Picea engelmannii (Burns and Honkala 1990).
Photos at right show meadow and timberline tree clumps. Such clumps are the common feature of a phytogeographic type, the forest-tundra parkland. Many subalpine species occur in forest-tundra parkland, but it is an especially common setting for subalpine fir (both var. bifolia and var. lasiocarpa). Typically such parklands form in settings where heavy winter snowfall shortens the growing season and lowers soil temperature to the point where tree seedling establishment is impeded. Isolated trees may establish in such settings during intervals of warmer and/or drier climate, or on places where topography or wind reduces snow depth, or sometimes they represent trees of other species. Solar radiation reflected from the isolated trees causes the snow around them to melt earlier in the season, creating a microsite with marginally better seedling establishment conditions. Over time a forest of seedlings grows up around the pioneer tree, forming a tree clump. Such clumps may persist as discrete units on the landscape for a thousand years or more until eradicated by fire or disease, or until climate and microsite changes transform the site into forest&emdash;or tundra. A variant of this process, meadow invasion, is also shown at right; it occurs when environmental change allows seedlings to establish in a meadow where they were previously excluded. Meadow invasion can have many causes, related to factors such as grazing, fire, and climate variation. See also Arno and Hammerly (1984).
Remarkable Specimens
Largest: Using the "points" system to combine diameter, dbh and crown spread, the official U.S. national champion has diameter 204 cm, height 38.1 m, crown spread 8 m in 1992, located at Cream Lake in Olympic National Park, WA (Van Pelt 1996); it is var. lasiocarpa. Oddly enough this tree was measured when Steve Arno found it in 1962, and was then remeasured in 1992 and in 2021, and all measurements are the same. In other words, it appears to have not grown, and to have remained the largest known specimen, for over 60 years. However, using estimated stem volume, the largest known tree was measured in 1993 at Wells Valley, also in Olympic National Park, and was then 183.5 cm dbh and 37.5 m tall, with a 7.9 m crown spread (Van Pelt email 2024.11.17). This tree's current status is unknown, but 2024 aerial photos suggest the area remains undisturbed.
Tallest: Diameter 107 cm, height 52 m, crown spread 7 m in 1988; on the Icicle Creek Trail in the Alpine Lakes Wilderness of WA (Van Pelt 1996). Variety not known, both bifolia and lasiocarpa occur in this area. This tree's current status is also unknown; Icicle Creek has experienced some severe fires in recent years, and the tree may be lost.
Var. arizonica: The largest is 33.8 m tall, 133 cm dbh, near Ruidoso, New Mexico. This is probably a pre-1996 measurement, and further details are unknown.
Var. bifolia: The tallest and largest was 37.2 m tall and 80 cm dbh, with a 5.9 m crown spread, in the San Juan Mountains of Colorado (Markworth 2013). This area has also experienced recent severe fires and the tree's current status is unknown.
Oldest: A tree-ring chronology covering the period 1496-2002 (crossdated after 1620) was collected at a site in southern Yukon, Canada by Mike Kenigsburg and Brian Luckman (doi.org/10.25921/64dh-1048). The oldest tree in the study provided a 507-year record. This age was nearly equalled by a tree in Yukon Territory with a crossdated age of 501 rings, reported by Luckman (2003; cited by RMTRR 2006). Based on its location, this tree was probably var. lasiocarpa. I sampled one tree of var. bifolia in the Medicine Bow Mountains, Wyoming, aged 494 years with over 90% of the age verified by cross-dating (Earle 1993). I have sampled (in 1986) trees in the Olympic Mountains of Washington with ring-counted ages of more than 450 years.
Ethnobotany
Due to high elevations and slow growth rates, subalpine fir is not widely used for timber, but when used, it provides lumber in building construction, crates, sashes, doors, frames, food containers, and pulp (Alexander et al. 1990). It may have greater economic significance for its contributions to watershed function, wildlife habitat, and recreational activities. Once, in 2008, it served as the U.S. Capitol Christmas tree.
Numerous studies of forest age structure have been conducted, some of which (e.g., Earle [1993]) have used dendrochronological methods.
Observations
I have seen var. lasiocarpa at the Mt. Washington ski area on Vancouver Island, British Columbia, where it is a very common subalpine tree in Strathcona Provinical Park. It is also in Olympic and Mount Rainier National Park, Washington, having there been the subject of numerous ecological studies of the trees' population dynamics, response to fire, habitat value, etc. (see notes and citations in Arno and Hammerly [1984]).
Var. bifolia is very common in the subalpine zone of mountains throughout its range. Among other places, you can find it easily in Glacier, Yellowstone, Grand Teton, and Rocky Mountain National Parks.
Var. arizonica is said to be common in the Sangre de Cristo and San Juan Mountains, and at Wolf Creek Pass, all locations in Colorado; and on the San Francisco Peaks in Arizona.
Remarks
Subalpine fir has weak apical dominance. Most conifers have strong apical dominance, meaning that the terminal bud (usually, the highest growing tip on the plant) produces indole acetic acid (IAA), a growth substance that inhibits development of apical dominance on any of the lower branches. This is why most conifers have a single erect trunk with a single live top. Subalpine fir, on the other hand, will commonly develop multiple points of apical dominance, usually in one of two situations, both illustrated at right. In one, an injury to the growing tip stops IAA production and more than one secondary growing tip independently assumes dominance. In the second, a lower branch is pressed to the ground and roots (this is called layering), after which the rooted branch assumes apical dominance independent of the mother tree. This tendency toward layering allows A. lasiocarpa to reproduce in situations, such as alpine settings, where the growing season is too short for normal cone and seed maturation to occur. Besides Abies, weak apical dominance is also common in many genera of the Cupressaceae, e.g. Sequoia and Thuja. Some conifers, mainly in the Cupressaceae, have weak apical dominance that results in a prostrate or decumbent growth habit.
Citations
Adams, R. P., C. J. Earle, and D. Thornburg. 2011. Taxonomy of infraspecific taxa of Abies lasiocarpa: leaf essential oils and DNA. Phytologia 93(1):87-101. Available at www.phytologia.org/.
Alexander, Robert R., Raymond C. Shearer, and Wayne D. Shepperd. 1990. Abies lasiocarpa (Hook.) Nutt. Subapline [sic] Fir, pp. 60-70 in Burns and Honkala (1990).
Arno, Stephen F. and Ramona Hammerly. 1984. Timberline: Mountain and Arctic Forest Frontiers. Seattle: The Mountaineers.
Harris, A. S. 1965. Subalpine fir [Abies lasiocarpa (Hook.) Nutt.] on Harris Ridge near Hollis, Prince of Wales Island, Alaska. Northwest Science 39:123-128.
Heusser, C. J. 1954. Alpine fir at Taku glacier, Alaska, with notes on its post glacial migration to the territory. Bulletin of the Torrey Botanical Club 81:83-86.
Luckman, B. H. 2003. Assessment of present, past and future climate variability in the Americas from treeline environments. IAI CRN03 Annual Report 2003.
Markworth, Matt. 2013.08.03. Re: Iowa Big Tree Guy Conquers Colorado. www.ents-bbs.org/viewtopic.php?f=70&t=5611#p24822, accessed 2014.08.17.
Silba, J. 2008. Journal of the International Conifer Preservation Society 15(2):42.
See also
Elwes and Henry 1906-1913 at the Biodiversity Heritage Library (Photo). This series of volumes, privately printed, provides some of the most engaging descriptions of conifers ever published. Although they only treat species cultivated in the U.K. and Ireland, and the taxonomy is a bit dated, still these accounts are thorough, treating such topics as species description, range, varieties, exceptionally old or tall specimens, remarkable trees, and cultivation. Despite being over a century old, they are generally accurate, and are illustrated with some remarkable photographs and lithographs.
Farjon, Aljos. 1990. Pinaceae: drawings and descriptions of the genera Abies, Cedrus, Pseudolarix, Keteleeria, Nothotsuga, Tsuga, Cathaya, Pseudotsuga, Larix and Picea. Königstein: Koeltz Scientific Books.
- Provides a detailed account, with illustrations.
Lanner (1983).
Lanner (1999).
MacKinnon et al. 1992.
FEIS database.
Zavarin, E., K. Snajberk, T. Reichert, and E. Tsien. 1970. On the geographic variability of the monoterpenes from the cortical blister oleoresin of Abies lasiocarpa. Phytochemistry 9:377-395.