Conservation Status

Juniperus convallium
Rehder et E.H. Wilson ex Sargent 1916
Common names
命枝圆柏 mizhi yuanbai.
Taxonomic notes
Syn: Sabina convallium (Rehder et E.H. Wilson) W.C. Cheng et L.K. Fu 1978; J. mekongensis Kom. 1924; J. ramulosa Florin 1927 (Farjon 2005).
This is a generally-recognized species regarded as sister to J. fargesii (various authorities) or J. carinata (Adams 2014). Farjon (2010) and Fu et al. (1999) recognize two varieties, convallium and Juniperus convallium var. microsperma (W.C. Cheng et L.K. Fu) Silba 1984 (syn: Sabina convallium var. microsperma W.C. Cheng et L.K. Fu 1975; Sabina microsperma (W.C. Cheng et L.K. Fu) W.C. Cheng et L.K. Fu 1983; J. microsperma (W.C. Cheng et L.K. Fu) R.P. Adams 2000), though Farjon acknowledges that the distinctiveness of var. microsperma is not well supported. Adams (2014) recognizes a forma, J. convallium f. pendula (Cheng et L.K. Fu) R.P Adams 1994 (syn: J. prezewalskii f. pendula R.P. Adams et G.L. Chu; Sabina przewalskii (Kom.) W.C. Cheng et L.K. Fu f. pendula W.C. Cheng et L.K. Fu 1975), noting that molecular analysis places it within J. convallium rather than J. przewalskii.
Gene-flow work comparing J. convallium with three other junipers of the Qinghai-Tibetan Plateau, J. tibetica, J. saltuaria and J. przewalskii, has given us a glimpse of the complex evolutionary history of the turbinate-cone juniper clade, finding evidence of "both incomplete lineage sorting and gene flow after species divergence" (Li et al. 2011; see this source for considerable further insights regarding this group).
Type: Western Sichuan, without precise locality, arid places, elev. 2500 m, August 1904, Veitch Expedition no. 3010 (Rehder and Wilson 1916).
Description
Dioecious or monoecious trees (rarely shrubs) to 20 m tall and 50 cm dbh, multi-stemmed or monopodial, with spreading or ascending branches forming a dense, rounded crown. Bark on larger stems gray-brown, exfoliating in longitudinal strips. Foliage branches densely arranged, short and stiff or longer and more lax. Foliar twigs slender, subterete or weakly quadrangular, 0.8-1.2 mm thick, covered with closely appressed leaves. Leaves on mature plants scale-like, decussate (sometimes ternate), imbricate, decurrent, 1.5-2 × 0.8-1 mm, rhombic, obtuse; epistomatic, with stomata in a basally divided band; abaxially glandular (but inconspicuous in var. microsperma); gland concave or convex, yellowish or darker than the light green or gray-green leaf. Pollen cones numerous, solitary, terminal on short twigs, subglobose, 2-3 mm long; microsporophylls decussate, 6-8, peltate-triangular, with obtuse apex, bearing 2-3 abaxial pollen sacs near the lower margin. Seed cones terminal on short, curved or erect branchlets, maturing in the second season to subglobose or ovoid-conical, 5-8 × 5-6 mm, light reddish brown to purplish black, sometimes glaucous. Bract-scale complexes 4-6, decussate, entirely fused, sometimes sutures faintly visible near apex of cone; bract tip usually hidden or barely visible; scale tissue dry pulpy or more succulent. Seeds 1 per cone, subglobose to ovoid-conical, 3-5 mm across, shallowly grooved or bifacially ridged, with small resin pits near base (Farjon 2010). See García Esteban et al. (2004) for a detailed characterization of the wood anatomy.
Var. microsperma differs in having leaves inconspicuously glandular, keeled near apex; with ovoid seed cones 5-7 mm on the long axis; seeds slightly flattened (Farjon 2010). F. pendula simply has drooping foliage (Adams 2014).
"[R]eadily distinguished by its shining, chestnut-brown, decurved one-seeded fruit. In habit
and appearance it somewhat resembles J. excelsa Bieberstein, but this has much larger, glaucous-black, 2-5-seeded, erect fruit. It may also be compared with J. saltuaria Rehder & Wilson which has more or less 4-sided branchlets, slightly keeled leaves with inconspicuous dorsal gland and small, erect, jet-black fruit. Juniperus convallium is apparently a rare tree and, as far as our knowledge goes, is confined to the warm and more arid parts of the principal river valleys of the Chino-Thibetan borderland. The tree is very Cypress-like in appearance and is frequently very glaucous" (Rehder and Wilson 2016).
Distribution and Ecology
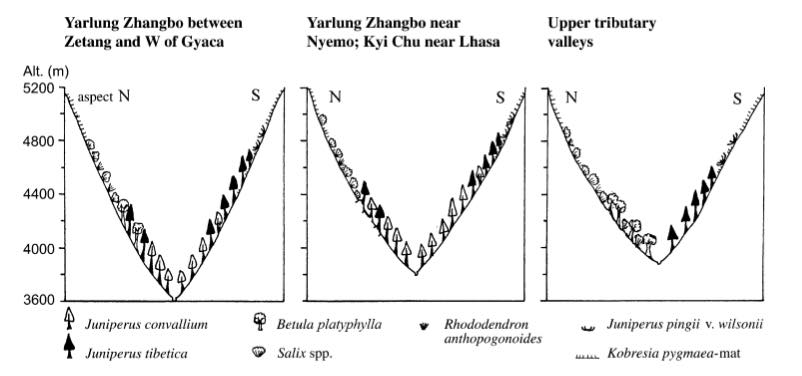
China: S Gansu, SE Qinghai, W Sichuan, E Xizang (Tibet), and NW Yunnan at 2200-4430 m elevation. Occurs in and marginal to areas of monsoonal climate, thus mainly summer rainfall; distribution indicates rainfall of at least 300 mm per year. Found on both carbonate and silicate substrates in high montane to subalpine coniferous forest, juniper woodland, and occasionally in the ecotone to alpine steppe (see illustration). Associates include Abies, Picea, Juniperus saltuaria, J. tibetica, Quercus aquifolioides, Seriphidium (Artemisia), Caragana, and Rosa; the woodlands are often degraded to scrub-pasture. Low winter temperatures, to which this species is more sensitive than J. tibetica, restrict it to lower elevations in many areas (cold air drainage effect?), but there are mixed stands especially on S-SW exposed slopes where J. convallium can occur at higher altitudes (Farjon 2010). Hardy to Zone 7 (cold hardiness limit between -17.7°C and -12.2°C) (Bannister and Neuner 2001).
Miehe et al. (2007) present evidence that the extant distribution of this species is relict, reflecting environmental changes with both climatic and anthropogenic components. In their study, an inventory of isolated tree stands surrounded by desert pastures in southern Tibet revealed more than 50 sites with vigorous trees of Juniperus convallium and J. tibetica. The trees are religious landmarks of the Tibetan Buddhists, a circumstance that has led to fairly good records of these locations. Vegetation records, rainfall correlations and data from local climate stations indicate that forest relicts fragmented through human activity could regenerate if current grazing and deforestation practices were halted. A first pollen diagram from Lhasa shows forest decline associated with the presence of humans since at least 4,600 yr BP, with the currently degraded commons developed in the last 600 yr. The westernmost J. convallium occurring in these relict stands are in the valley of of the Yarlung Zhangbo, the remains of a stand largely destroyed during the Cultural Revolution, longitude 88.7° E.
Remarkable Specimens
No data as of 2023.03.03.
Ethnobotany
Observations
Remarks
Farjon (2010) proposes that the epithet derives from Latin convallis, a deep, enclosed valley, in reference to its habitat.
Citations
Adams, R. P. 2014. Junipers of the World: The Genus Juniperus, 4th ed. Bloomington, IN: Trafford Publ. Co.
Farjon, Aljos. 2010. A Handbook of the World's Conifers. Leiden, Netherlands: Brill Academic Publishers.
Li Zhonghu, Jiabin Zou, Kangshan Mao, Kao Lin, Haipeng Li, Jianquan Liu, Thomas Kallman, and Martin Lascoux. 2011. Population genetic evidence for complex evolutionary histories of four high altitude juniper species in the Qinghai–Tibetan Plateau. Evolution 66(3):831-845, doi:10.1111/j.1558-5646.2011.01466.x.
Miehe, G., S. Miehe, M. Will, L. Opgenoorth, L. Duo, T. Dorgeh, and J. Liu. 2007. An inventory of forest relicts in the pastures of Southern Tibet (Xizang A.R., China). Plant Ecology 194(2):157–177.
Opgenoorth, L., G. G. Vendramin, K. S. Mao, G. Miehe, S. Miehe, S. Liepelt,
J. Q. Liu, and B. Ziegenhagen. 2010. Tree endurance on the Tibetan plateau marks the world’s highest known tree line of the Last Glacial Maximum. New Phytol. 185:332–342.
Rehder and E. H. Wilson. 1916. Juniperus convallium. P. 62 in Sargent, C. S. (ed.), Plantae Wilsoniae, Publications of the Arnold Arboretum No. 4. Cambridge: University Press. Available at the Biodiversity Heritage Library, accessed 2020.01.29.
See also
Farjon (2005) provides a detailed account, with illustrations.
Urgenson, L., A. H. Schmidt, J. Combs, S. Harrell, T. Hinckley, Q. Yang, Z. Ma, L. Yongxian, L. Hongliang, and A. MacIver. 2014. Traditional livelihoods, conservation and meadow ecology in Jiuzhaigou National Park, Sichuan, China. Hum Ecol Interdiscip J. 42(3): 481–491, available ResearchGate, accessed 2020.01.29.